Flavínadeníndinukleotid
Flavínadeníndinukleotid (FAD) je redoxne aktívny koenzým spojený s rôznymi bielkovinami, ktorý sa podieľa na niekoľkých enzymatických reakciách v metabolizme. Flavoproteíny sú bielkoviny, ktoré obsahujú flavínovú skupinu, ktorá môže byť vo forme FAD alebo flavínmononukleotidu (FMN). Je známych veľa flavoproteínov, napríklad časti komplexu sukcinátdehydrogenázy, α-ketoglutarátdehydrogenáza alebo jedna z častí komplexu pyruvátdehydrogenázy.
| Flavínadeníndinukleotid | |
 | |
 | |
| Všeobecné vlastnosti | |
| Sumárny vzorec | C27H33N9O15P2 |
| Synonymá | FAD |
| Fyzikálne vlastnosti | |
| Molárna hmotnosť | 785,557 g/mol |
| pKA | 1,128 |
| pKB | 12,868 |
| Ďalšie informácie | |
| Číslo CAS | 146-14-5 |
| Pokiaľ je to možné a bežné, používame jednotky sústavy SI. Ak nie je hore uvedené inak, údaje sú za normálnych podmienok. | |
FAD sa môže vyskytovať v štyroch redoxných stavoch, ktorými sú flavín-N(5)-oxid, chinón, semichinón a hydrochinón.[1] FAD prechádza medzi týmito stavmi prijatím alebo darovaním elektrónov. FAD vo svojej úplne oxidovanej forme alebo vo forme chinónu prijíma dva elektróny a dva protóny (ióny H+), čím sa z neho stáva FADH2 (hydrochinónová forma). Semichinón (FADH•) môže vzniknúť buď redukciou FAD alebo oxidáciou FADH2, v oboch prípadoch prijatím alebo darovaním jedného elektrónu a jedného protónu. Niektoré proteíny ho však vytvárajú a udržiavajú v podobe flavín-N(5)-oxidu.[2][3]
História
upraviťFlavoproteíny boli prvýkrát objavené v roku 1879 oddelením zložiek kravského mlieka. Pre ich mliečny pôvod a žltý pigment sa spočiatku nazývali laktochrómy.[4] Trvalo 50 rokov, kým vedecká komunita urobila zásadný pokrok v identifikácii molekúl zodpovedných za žltý pigment. 30. roky 20. storočia odštartovali oblasť výskumu koenzýmov publikovaním mnohých štruktúr flavínových a nikotínamidových derivátov a ich významných úloh v redoxnej katalýze. Nemeckí vedci Otto Warburg a Walter Christian objavili v roku 1932 žltý proteín z kvasiniek potrebný na bunkové dýchanie. Ich kolega Hugo Theorell rozdelil tento žltý enzým na apoenzým a žltý pigment a ukázal, že ani enzým, ani pigment nie sú schopné samostatne oxidovať NADH, ale ich zmiešaním sa aktivita obnovila. Theorell v roku 1937 potvrdil, že pigmentom je fosfátový ester riboflavínu, flavínmononukleotid (FMN), čo bol prvý priamy dôkaz enzýmových kofaktorov.[5] Warburg a Christian potom podobnými experimentmi v roku 1938 zistili, že FAD je kofaktorom oxidázy D-aminokyselín.[6] Warburgova práca, ktorá prepojila nikotínamid s prenosom hydridu a objav flavínov, vydláždili cestu mnohým vedcom v 40. a 50. rokoch, kedy bolo objavené veľké množstvo redoxnej biochémie a tieto reakcie boli spojené v dráhach, ako sú citrátový cyklus a syntéza ATP.
Vlastnosti
upraviťFAD pozostáva z dvoch častí: adenínového nukleotidu (adenozínmonofosfátu) a flavínmononukleotidu (FMN), ktoré sú navzájom premostené prostredníctvom svojich fosfátových skupín. Adenín je naviazaný na cyklickú ribózu na 1' uhlíku, zatiaľ čo fosfát je naviazaný na ribózu na 5' uhlíku za vzniku adenínového nukleotidu. Riboflavín je tvorený väzbou uhlík-dusík (CN) medzi flavínom (modifikovaným izoaloxazínom) a ribitolom. Fosfátová skupina je potom naviazaná na koncový uhlík ribózy, čím vzniká FMN. Pretože väzba medzi izoaloxazínom a ribitolom sa nepovažuje za glykozidovú väzbu, flavínmononukleotid nie je skutočným nukleotidom.[7] Názov dinukleotid je teda zavádzajúci, avšak flavínmononukleotidová skupina je svojou štruktúrou a chemickými vlastnosťami stále veľmi blízka nukleotidu.

FAD možno redukovať na FADH2 pridaním 2 H+ a 2 e -. FADH2 sa potom môže oxidovať aj stratou 1 H + a 1 e − za vzniku FADH. Ďalšou stratou 1 H+ a 1 e- môže byť obnovený FAD. K tvorbe FAD môže dôjsť aj redukciou a dehydratáciou flavín-N(5)-oxidu.[8] Na základe oxidačného stavu získavajú flavíny vo vodnom roztoku špecifické farby. Flavín-N(5)-oxidová forma je žltooranžová, FAD (oxidovaná forma) je žltý, FADH (poloredukovaný) je buď modrý alebo červený podľa pH a FADH2 (úplne redukovaná forma) je bezfarebný.[9][10] Zmena formy môže mať veľký vplyv na ďalšie chemické vlastnosti. Napríklad FAD, plne oxidovaná forma, podlieha nukleofilnému ataku, zatiaľ čo plne redukovaná forma, FADH2, má vysokú polarizovateľnosť a polovične redukovaná forma, FADH, je vo vodnom roztoku nestabilná.[11] FAD je aromatický kruhový systém, zatiaľ čo FADH2 nie je.[12] To znamená, že FADH2 má výrazne vyššiu energiu bez stabilizácie prostredníctvom rezonancie, ktorú poskytuje aromatická štruktúra. FADH2 je teda molekula nesúca energiu, pretože po oxidácii znovu získava aromatickosť a uvoľňuje energiu reprezentovanú touto stabilizáciou.

Spektroskopické vlastnosti FAD a jeho variantov umožňujú sledovanie reakcií pomocou UV-Vis absorpčnej a fluorescenčnej spektroskopie. Každá forma FAD má odlišné absorpčné spektrum, čo uľahčuje pozorovanie zmien v oxidačnom stave.[11] Hlavné lokálne maximum absorpcie pre FAD je pozorované pri 450 nm s absorpčným koeficientom 11 300 M −1 cm −1.[13] Flavíny majú vo všeobecnosti fluorescenčnú aktivitu, keď nie sú viazané na bielkoviny (proteíny naviazané na deriváty flavínových nukleotidov sa nazývajú flavoproteíny). Túto vlastnosť možno využiť pri skúmaní proteínovej väzby, pretože možno sledovať stratu fluorescencie, keď sa flavíny naviažu na bielkovinu.[11] Oxidované flavíny majú vysoké absorbancie okolo 450 nm a fluoreskujú pri asi 515-520 nm.[9]
Chemické stavy
upraviťV biologických systémoch pôsobí FAD ako akceptor H+ a e− v jeho plne oxidovanej forme, ako akceptor alebo donor vo forme FADH a ako donor v redukovanej forme FADH2. Nižšie uvedený diagram sumarizuje redoxné zmeny, ktorými môže FAD prejsť.

Okrem toho môžu vznikať a reagovať ďalšie reaktívne formy FAD. Tieto reakcie zahŕňajú prenos elektrónov a vznik/zánik chemických väzieb. FAD je schopný prispievať k chemickým reakciám v rámci biologických systémov ako súčasť rôznych reakčných mechanizmov. Nasledujúce obrázky zobrazujú všeobecné podoby niektorých reakcií, do ktorých sa môže FAD zapojiť.
Mechanizmy 1 a 2 predstavujú príjem hydridu, pri ktorom molekula prijíma v procese jeden hydridový anión, a to v podobe vodíkového katiónu a dvoch elektrónov. Mechanizmy 3 a 4 predstavujú tvorbu radikálov a stratu hydridu. Radikály obsahujú atómy s nepárovými elektrónmi a sú veľmi (re)aktívne. Strata hydridu je opačný proces k vyššie popísanému príjmu hydridu. Mechanizmy 5 a 6 ukazujú nukleofilnú adíciu a reakciu využívajúcu uhlíkový radikál.
 |
 |
 |
 |
 |
 |
Biosyntéza
upraviťFAD hrá hlavnú úlohu ako enzýmový kofaktor spolu s flavínmononukleotidom (FMN), ďalšou molekulou odvodenou od riboflavínu.[8] Baktérie, huby a rastliny vedia produkovať riboflavín, ale iné eukaryoty, ako napríklad ľudia, stratili schopnosť ho syntetizovať.[9] Preto ľudia musia získavať riboflavín, tiež známy ako vitamín B2, zo zdrojov potravy.[14] Riboflavín sa vo všeobecnosti prijíma v tenkom čreve a potom sa presúva do buniek prostredníctvom nosných proteínov.[9] Riboflavínkináza (EC 2.7.1.26) pridáva fosfátovú skupinu k riboflavínu za vzniku FMN a potom naňho FAD syntetáza pripája adenínový nukleotid za vzniku FAD, pričom oba kroky vyžadujú ATP.[9] Baktérie majú vo všeobecnosti jeden bifunkčný enzým, ale archeóny a eukaryoty zvyčajne používajú dva odlišné enzýmy.[9] Súčasný výskum naznačuje, že v cytosóle a mitochondriách existujú odlišné izoformy.[9] Zdá sa, že FAD je syntetizovaný na oboch miestach a potenciálne transportovaný tam, kde je to potrebný.[11]

Funkcia
upraviťFlavoproteíny využívajú jedinečnú a všestrannú štruktúru flavínových skupín na katalýzu náročných redoxných reakcií. Keďže flavíny majú viacero redoxných stavov, môžu sa podieľať na procesoch, ktoré zahŕňajú prenos jedného alebo dvoch elektrónov, atómov vodíka alebo iónov H3O+. N5 a C4a plne oxidovaného flavínového kruhu sú tiež náchylné na nukleofilný útok.[15] Túto širokú škálu stavov ionizácie a modifikácie flavínovej časti možno pripísať izoaloxazínovému kruhovému systému a schopnosti flavoproteínov drasticky narušiť kinetické parametre flavínov, vrátane flavínadeníndinukleotidu (FAD), po naviazaní.
Počet flavoproteínov kódovaných v genóme (flavoproteóm) je závislý na konkrétnom druhu a môže sa pohybovať od 0,1 % do 3,5 %, pričom ľudia majú 90 génov, ktoré kódujú flavoproteíny.[16] FAD je komplexnejšia a hojnejšia forma flavínu a uvádza sa, že sa viaže na 75 % celkového flavoproteómu[16] a 84 % ľudských kódovaných flavoproteínov.[17] Bunkové koncentrácie voľných alebo nekovalentne viazaných flavínov v rôznych kultivovaných cicavčích bunkových líniách boli stanovené pre FAD (2,2-17,0 amol/bunka) a FMN (0,46-3,4 amol/bunka).[18]
FAD má pozitívnejší redukčný potenciál ako NAD+ a je to veľmi silné oxidačné činidlo. Bunky ho využívajú pri mnohých energeticky náročných oxidačných reakciách, napríklad dehydrogenácii C–C väzby za vzniku alkénu. Proteíny využívajúce FAD fungujú vo veľkom množstve metabolických dráh vrátane transportu elektrónov, opravy DNA, biosyntézy nukleotidov, beta-oxidácie mastných kyselín, katabolizmu aminokyselín, ako aj syntézy ďalších kofaktorov, ako sú CoA, CoQ a hemových skupín. Jedna zo známejších reakcií je súčasťou citrátového cyklu – sukcinátdehydrogenáza (komplex II v elektrónovom transportnom reťazci) vyžaduje kovalentne naviazaný FAD, aby mohla katalyzovať oxidáciu sukcinátu na fumarát, ktorá je spriahnutá s redukciou ubichinónu na ubichinol.[11] Vysokoenergetické elektróny z tejto oxidácie sa na chvíľu uložia do redukovaného FADH2. FADH2 sa potom vráti do podoby FAD a presúva svoje dva vysokoenergetické elektróny cez elektrónový transportný reťazec, pričom energia uložená v FADH2 postačuje na výrobu 1,5 ekvivalentu ATP[19] oxidačnou fosforyláciou. Niektoré redoxné flavoproteíny viažu FAD nekovalentne, ako napríklad acetyl-CoA-dehydrogenázy, ktoré sa podieľajú na beta-oxidácii mastných kyselín a katabolizme aminokyselín, ako sú leucín (izovaleryl-CoA dehydrogenáza), izoleucín (dehydrogenáza acyl-CoA s krátkym/rozvetveným reťazcom), valín (izobutyryl-CoA dehydrogenáza) a lyzín (glutaryl-CoA dehydrogenáza).[20] Ďalšími príkladmi enzýmov využívajúcich FAD, ktoré regulujú metabolizmus, sú glycerol-3-fosfátdehydrogenáza (syntéza triglyceridov) a xantínoxidáza, ktorá sa podieľa na katabolizme purínových nukleotidov.[21] Nekatalytické funkcie, ktoré môže hrať FAD vo flavoproteínoch, zahŕňajú štruktúrne úlohy alebo sa podieľa na funkcii fotoreceptorov citlivých na modré svetlo, ktoré regulujú biologické hodiny a vývoj a generovanie svetla v bioluminiscenčných baktériách.[20]
Flavoproteíny
upraviťFlavoproteíny majú ako prostetickú skupinu molekulu FMN alebo FAD. Táto prostetická skupina môže byť viazaná kovalentne alebo nekovalentne. Len asi 5-10% flavoproteínov má FAD viazaný kovalentne, ale tieto enzýmy majú väčšiu redoxnú silu.[11] V niektorých prípadoch môže FAD poskytnúť štruktúrnu oporu pre aktívne miesta enzýmov alebo poskytnúť stabilizáciu medziproduktov počas katalýzy.[20] Na základe dostupných štruktúrnych údajov možno známe miesta viažuce FAD rozdeliť na viac ako 200 typov.[22]
V ľudskom genóme je kódovaných 90 flavoproteínov; asi 84 % vyžaduje FAD a zvyšných asi 16 % vyžaduje FMN, zatiaľ čo 5 proteínov vyžaduje prítomnosť oboch.[17] Flavoproteíny sa nachádzajú hlavne v mitochondriách kvôli ich redoxnej sile.[17] Spomedzi všetkých flavoproteínov, 90 % katalyzuje redoxné reakcie a zvyšných 10 % sú transferázy, lyázy, izomerázy a ligázy.[16]
Oxidácia väzieb uhlík-heteroatóm
upraviťUhlík-dusík
upraviťMonoaminooxidáza (MAO) je rozsiahle študovaný flavoenzým kvôli jeho biologickému významu pri katabolizme norepinefrínu, serotonínu a dopamínu. MAO oxiduje primárne, sekundárne a terciárne amíny, ktoré potom neenzymaticky hydrolyzujú z imínu na aldehyd alebo ketón. Aj keď táto trieda enzýmov bola dôkladne študovaná, o mechanizme jej účinku sa stále diskutuje. Boli navrhnuté dva mechanizmy: radikálny mechanizmus a nukleofilný mechanizmus. Radikálový mechanizmus je všeobecne menej prijatý, pretože neexistuje žiadny spektrálny dôkaz alebo dôkaz z elektrónovej paramagnetickej rezonancie, ktorý by ukazoval prítomnosť radikálového medziproduktu. Nukleofilný mechanizmus je uprednostňovaný, pretože je podporovaný štúdiami miestne cielenej mutagenézy, ktoré mutovali dva tyrozínové zvyšky, u ktorých sa očakávalo, že zvýšia nukleofilitu substrátov.[23]

Uhlík-kyslík
upraviťGlukózaoxidáza (GOX) katalyzuje oxidáciu β-D-glukózy na D-glukono-δ-laktón so súčasnou redukciou flavínu viazaného na enzým. GOX existuje ako homodimér, pričom každá podjednotka viaže jednu molekulu FAD. Kryštálové štruktúry ukazujú, že FAD sa viaže hlboko vnútri enzýmu blízko rozhrania diméru. Štúdie takisto ukázali, že po nahradení FAD 8-hydroxy-5-karba-5-deaza-FAD sa stereochémia reakcie je daná reakciou s re stranou flavínu. Počas premeny boli pozorované neutrálne a aniónové semichinóny, čo naznačuje radikálny mechanizmus.[23]

Uhlík-síra
upraviťPrenylcysteínlyáza katalyzuje štiepenie prenylcysteínu (proteínovej modifikácie) za vzniku izoprenoidového aldehydu a uvoľneného cysteínového zvyšku na cielenom proteíne. FAD sa viaže na prenylcysteínlyázu nekovalentne. Neuskutočnilo sa veľa mechanistických štúdií zameraných na premeny flavínu, ale navrhovaný mechanizmus je uvedený nižšie. Predpokladá sa prenos hydridu z uhlíkového atómu C1 prenylovej skupiny na FAD, čo vedie k redukcii flavínu na FADH2. Zároveň vzniká karbokatión, ktorý je stabilizovaný susedným atómom síry. FADH2 potom reaguje s molekulárnym kyslíkom, aby sa obnovil oxidovaný enzým.[23]

Uhlík-uhlík
upraviťUDP-N-acetylenolpyruvylglukózamínreduktáza (MurB) je enzým, ktorý katalyzuje NADPH-dependentnú redukciu enolpyruvyl-UDP-N-acetylglukózamínu (substrátu) na zodpovedajúcu D-laktylovú zlúčeninu kyselinu UDP-N-acetylmuramovú (produkt). MurB je monomér a obsahuje jednu molekulu FAD. Než sa substrát premení na produkt, najprv musí NADPH redukovať FAD. Akonáhle sa NADP+ disociuje, môže sa naviazať substrát a redukovaný flavín môže redukovať produkt.[23]

Premeny tiol/disulfid
upraviťGlutatiónreduktáza (GR) katalyzuje redukciu glutatióndisulfidu (GSSG) na glutatión (GSH). GR vyžaduje FAD a NADPH na sprostredkovanie tejto reakcie – najprv sa musí previesť hydrid z NADPH na FAD. Redukovaný flavín potom môže pôsobiť ako nukleofil na napadnutie disulfidu, čím sa vytvorí adukt C4a-cysteín. Eliminácia tohto aduktu vedie k prenosu náboja v komplexe flavín-tiolát.[23]

Reakcie s prenosom elektrónov
upraviťEnzýmy typu cytochrómu P450, ktoré katalyzujú monooxygenázové (hydroxylačné) reakcie, využívajú prenos dvoch elektrónov z FAD na P450. V eukaryotoch sa nachádzajú dva typy systémov P450.
Systémy P450, ktoré sa nachádzajú v endoplazmatickom retikule, využívajú cytochróm P450 reduktázu, ktorá obsahuje FAD aj FMN. Dva elektróny na redukovanom FAD (teda FADH2) sa jeden po druhom prenesú do FMN a potom sa jeden elektrón prenesie z FMN na hem, ktorý sa nachádza v P450.[24]
Systémy P450, ktoré sa nachádzajú v mitochondriách, vyžadujú dva proteíny na prenos elektrónov: adrenodoxínreduktázu (AR), ktorá obsahuje FAD, a adrenodoxín, proteín obsahujúci malú železo-sírnu skupinu. FAD je zabudovaný do domény AR, ktorá viaže FAD.[25][26] FAD v AR je redukovaný na FADH2 prenosom dvoch elektrónov z NADPH, ktorý sa viaže v NADPH viažucej doméne AR. Štruktúra tohto enzýmu je vo veľkej miere zachovávaná, aby sa zachovalo presné usporiadanie donoru elektrónov NADPH a akceptoru FAD pre účinný prenos elektrónov.[26] Dva elektróny v redukovanom FAD sa jeden po druhom prenesú na adrenodoxín, ktorý zase prenesie jeden elektrón na hemovú skupinu mitochondriálneho P450.[27]
Štruktúry reduktáz mikrozomálnych a reduktáz mitochondriálnych systémov P450 sú úplne odlišné a nevykazujú žiadnu homológiu.[24]
Redoxné reakcie
upraviťp-hydroxybenzoáthydroxyláza (PHBH) katalyzuje oxygenáciu p-hydroxybenzoátu pOHB) na 3,4-dihydroxybenzoát (3,4-diOHB). Na túto reakciu sú potrebné FAD, NADPH a molekulárny kyslík. NADPH najprv prenesie ekvivalent hydridu na FAD, čím vzniká FADH− a potom NADP+ z enzýmu disociuje. Redukovaná PHBH potom reaguje s molekulárnym kyslíkom za vzniku flavín-C(4a)-hydroperoxidu. Flavínhydroperoxid rýchlo hydroxyluje pOHB a potom štiepi vodu, čím sa regeneruje oxidovaný flavín.[23] Alternatívny mechanizmus oxygenácie sprostredkovaný flavínom spočíva v použití flavín-N(5)-oxidu namiesto flavín-C(4a)-(hydro)peroxidu.[2][3]

Neredoxné reakcie
upraviťChorizmátsyntáza (CS) katalyzuje posledný krok v šikimátovej dráhe — tvorbu chorizmátu. Sú známe dve triedy CS, pričom obe vyžadujú redukovaný flavín (v podobe FMN alebo FAD[28]), ale líšia sa v potrebe NADPH ako redukčného činidla. Navrhovaný mechanizmus pre CS zahŕňa prítomnosť radikálov. Radikál flavínu nebol detegovaný spektroskopicky bez použitia analógu substrátu, čo naznačuje, že existuje len veľmi krátku dobu. Pri použití fluórovaného substrátu bol však detegovaný neutrálny flavínový semichinón.[23]

Komplexné flavoenzýmy
upraviťGlutamátsyntáza katalyzuje premenu 2-oxoglutarátu na L-glutamát, pričom ako zdroj dusíka pre reakciu slúži L-glutamín. Všetky glutamátsyntázy sú železo-sírne flavoproteíny obsahujúce železo-sírny klaster a FMN i FAD. Glutamátsyntázy sú kategorizované do troch tried na základe svojich sekvencií a biochemických vlastností. Aj keď existujú tri triedy tohto enzýmu, predpokladá sa, že všetky pôsobia rovnakým mechanizmom a líšia sa len tým, čo najskôr redukuje FMN. Enzým produkuje dve molekuly glutamátu: jednu hydrolýzou glutamínu (vzniká glutamát a amoniak) a druhú, ktorá vzniká atakom amoniaku produkovaným v prvej reakcii na 2-oxoglutarát, ktorý je redukovaný FMN na glutamát.[23]
Klinický význam
upraviťOchorenia súvisiace s flavoproteínmi
upraviťVzhľadom na dôležitosť flavoproteínov nie je prekvapujúce, že približne 60 % ľudských flavoproteínov spôsobuje ľudské ochorenia, ak sú v nich prítomné mutácie.[17] V niektorých prípadoch je to spôsobené zníženou afinitou k FAD alebo FMN, v takých prípadoch potom môže nadmerný príjem riboflavínu zmierniť symptómy ochorenia, napríklad u mnohonásobného deficitu acyl-CoA dehydrogenázy.[9] Okrem toho môže samotný nedostatok riboflavínu (a výsledný nedostatok FAD a FMN) spôsobiť zdravotné problémy.[9] Napríklad u pacientov s ALS sú znížené hladiny syntézy FAD.[9] Obe tieto cesty môžu viesť k rôznym symptómom, vrátane vývojových alebo gastrointestinálnych abnormalít, chybného odbúravania tuku, anémie, neurologických problémov, rakoviny alebo srdcových chorôb, migrény, zhoršeného videnia a kožných lézií.[9] Farmaceutický priemysel preto vyrába riboflavín na doplnenie stravy pre podobné prípady. V roku 2008 bola celosvetová potreba riboflavínu 6 000 ton ročne s výrobnou kapacitou 10 000 ton.[4] Tento trh, ktorého hodnota predstavuje 150 až 500 miliónov amerických dolárov, nie je len pre medicínske aplikácie, ale súvisí i s použitím riboflavínu ako doplnku ku krmivu pre zvieratá v poľnohospodárskom priemysle a ako potravinárske farbivo.[4]
Vývoj liekov
upraviťDizajn nových antibakteriálnych liekov má význam vo vedeckom výskume i naďalej, pretože rezistencia baktérií na bežné antibiotiká sa zvyšuje. Špecifický metabolický proteín, ktorý využíva FAD (komplex II) je životne dôležitý pre bakteriálnu virulenciu, a preto zacielenie syntézy FAD alebo vytváranie analógov FAD by mohlo byť užitočnou oblasťou výskumu.[29] Vedci už určili dve štruktúry, ktoré FAD zvyčajne nadobúda, keď sa naviaže na bielkovinu: buď „natiahnutú“, kedy sú adenínová a izoaloxazínová skupina ďaleko od seba, alebo „motýľovú“ konformáciu, v ktorej sa molekula v podstate zloží na polovicu, čo vedie k stohovaniu adenínového a izoaloxazínového kruhu.[14] Imitátory FAD, ktoré sú schopné viazať sa podobným spôsobom, ale neumožňujú fungovanie proteínu, by mohli byť užitočnými mechanizmami inhibície bakteriálnej infekcie.[14] Alternatívne môžu lieky blokujúce syntézu FAD dosiahnuť rovnaký cieľ – tento spôsob je obzvlášť zaujímavý, pretože ľudská a bakteriálna syntéza FAD sa spolieha na veľmi odlišné enzýmy, čo znamená, že liek cielený na bakteriálnu FAD syntázu by pravdepodobne neinterferoval s ľudskými enzýmami FAD syntázy.[30]
Optogenetika
upraviťOptogenetika umožňuje riadenie biologických dejov neinvazívnym spôsobom.[31] Táto oblasť v posledných rokoch pokročila s množstvom nových nástrojov, vrátane tých, ktoré spúšťajú citlivosť na svetlo, ako sú domény FAD využívajúce modré svetlo (BLUF, z anglického blue-light-utilizing FAD domains). BLUF sa skladajú zo sekvencie s dĺžkou 100 až 140 aminokyselín, ktorá bola odvodená z fotoreceptorov v rastlinách a baktériách.[31] Podobne ako pri iných fotoreceptoroch spôsobuje svetlo štruktúrne zmeny v doméne BLUF, čo vedie k narušeniu následných interakcií.[31] Súčasný výskum skúma proteíny s pripojenou doménou BLUF a to, ako môžu rôzne vonkajšie faktory ovplyvniť tieto proteíny.[31]
Monitorovanie liečby
upraviťV tele je množstvo molekúl, ktoré majú prirodzenú fluorescenciu, vrátane tryptofánu, kolagénu, FAD, NADH a porfyrínov.[32] Vedci to využili na monitorovanie priebehu ochorenia alebo účinnosti liečby či na pomoc pri diagnostike. Napríklad natívna fluorescencia FAD a NADH sa líši v normálnom tkanive a orálnej submukóznej fibróze, čo je skorý príznak invazívnej rakoviny úst.[32] Lekári preto používajú fluorescenciu na pomoc pri diagnostike a monitorovaní liečby na rozdiel od štandardnej biopsie.[32]
Galéria
upraviť-

-
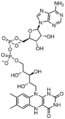 Redukovaný FAD (FADH2)
Redukovaný FAD (FADH2)


Referencie
upraviť- ↑ Unusual flavoenzyme catalysis in marine bacteria. Current Opinion in Chemical Biology, 2016-04-01, s. 31–39. ISSN 1879-0402. DOI: 10.1016/j.cbpa.2016.01.001. PMID 26803009.
- ↑ a b Flavin-mediated dual oxidation controls an enzymatic Favorskii-type rearrangement.. Nature, 28 November 2013, s. 552–6. DOI: 10.1038/nature12643. PMID 24162851.
- ↑ a b Biochemical Establishment and Characterization of EncM's Flavin-N5-oxide Cofactor. Journal of the American Chemical Society, 2015-07-01, s. 8078–8085. ISSN 1520-5126. DOI: 10.1021/jacs.5b03983. PMID 26067765.
- ↑ a b c Genetic control of biosynthesis and transport of riboflavin and flavin nucleotides and construction of robust biotechnological producers. Microbiology and Molecular Biology Reviews, Jun 2011, s. 321–60. DOI: 10.1128/mmbr.00030-10. PMID 21646432.
- ↑ HAYASHI, Hideyuki. B Vitamins and Folate: Chemistry, Analysis, Function and Effects. Cambridge, UK : The Royal Society of Chemistry, 2013. ISBN 978-1-84973-369-4. S. 7.
- ↑ Isolation of the prosthetic group of the amino acid oxidase. Biochemische Zeitschrift, 1938, s. 150–168.
- ↑ METZLER, David E.; METZLER, Carol M.; SAUKE, David J.. Biochemistry. 2nd. vyd. San Diego : Harcourt, Academic Press, 2003. Dostupné online. ISBN 978-0-12-492541-0.
- ↑ a b DEVLIN, Thomas M. Textbook of Biochemistry: with Clinical Correlations. 7th. vyd. Hoboken, NJ : John Wiley & Sons, 2011. ISBN 978-0-470-28173-4.
- ↑ a b c d e f g h i j k Biosynthesis of flavin cofactors in man: implications in health and disease. Current Pharmaceutical Design, 2013, s. 2649–75. DOI: 10.2174/1381612811319140014. PMID 23116402.
- ↑ Flavin-mediated dual oxidation controls an enzymatic Favorskii-type rearrangement. Nature, 2013-11-28, s. 552–556. ISSN 1476-4687. DOI: 10.1038/nature12643. PMID 24162851.
- ↑ a b c d e f Emerging concepts in the flavinylation of succinate dehydrogenase. Biochimica et Biophysica Acta (BBA) - Bioenergetics, May 2013, s. 627–36. DOI: 10.1016/j.bbabio.2013.01.012. PMID 23380393.
- ↑ LIU, Shijie. Bioprocess Engineering: Kinetics, Sustainability, and Reactor Design. [s.l.] : Newnes, 2012. Dostupné online. ISBN 978-0-444-63783-3.
- ↑ The FAD-dependent tricarballylate dehydrogenase (TcuA) enzyme of Salmonella enterica converts tricarballylate into cis-aconitate. Journal of Bacteriology, Aug 2006, s. 5479–86. DOI: 10.1128/jb.00514-06. PMID 16855237.
- ↑ a b c Conformational behavior of flavin adenine dinucleotide: conserved stereochemistry in bound and free states. The Journal of Physical Chemistry B, Nov 2014, s. 13486–97. DOI: 10.1021/jp507629n. PMID 25389798.
- ↑ MONTEIRA, Mariana. B Vitamins and Folate: Chemistry, Analysis, Function and Effects. Cambridge, UK : The Royal Society of Chemistry, 2013. ISBN 978-1-84973-369-4. S. 94.
- ↑ a b c Flavogenomics--a genomic and structural view of flavin-dependent proteins. The FEBS Journal, Aug 2011, s. 2625–34. DOI: 10.1111/j.1742-4658.2011.08202.x. PMID 21635694.
- ↑ a b c d The human flavoproteome. Archives of Biochemistry and Biophysics, Jul 2013, s. 150–62. DOI: 10.1016/j.abb.2013.02.015. PMID 23500531.
- ↑ Quantification of riboflavin, flavin mononucleotide, and flavin adenine dinucleotide in mammalian model cells by CE with LED-induced fluorescence detection. Electrophoresis, Feb 2015, s. 518–25. DOI: 10.1002/elps.201400451. PMID 25488801.
- ↑ STRYER, Lubert; BERG, Jeremy M.; TYMOCZKO, John L.. Biochemistry. 6th. vyd. New York : Freeman, 2007. Dostupné online. ISBN 978-0-7167-8724-2.
- ↑ a b c The diverse roles of flavin coenzymes--nature's most versatile thespians. The Journal of Organic Chemistry, Aug 2007, s. 6329–42. DOI: 10.1021/jo0703092. PMID 17580897.
- ↑ Vitamins, Minerals, Supplements [online]. 18 May 2020. Dostupné online.
- ↑ Structure-based classification of FAD binding sites: A comparative study of structural alignment tools. Proteins: Structure, Function, and Bioinformatics, 2016-11-01, s. 1728–1747. ISSN 1097-0134. DOI: 10.1002/prot.25158. PMID 27580869. (po anglicky)
- ↑ a b c d e f g h Flavin-Dependent Enzymes. Comprehensive Natural Products II Chemistry and Biology, 2010, s. 37–113.
- ↑ a b Hanukoglu I. Electron transfer proteins of cytochrome P450 systems. Adv. Mol. Cell Biol., 1996, s. 29–55. Dostupné online. DOI: 10.1016/S1569-2558(08)60339-2.
- ↑ The structure of adrenodoxin reductase of mitochondrial P450 systems: electron transfer for steroid biosynthesis. Journal of Molecular Biology, Jun 1999, s. 981–90. DOI: 10.1006/jmbi.1999.2807. PMID 10369776.
- ↑ a b Conservation of the Enzyme-Coenzyme Interfaces in FAD and NADP Binding Adrenodoxin Reductase-A Ubiquitous Enzyme. Journal of Molecular Evolution, 2017, s. 205–218. DOI: 10.1007/s00239-017-9821-9. PMID 29177972.
- ↑ Mitochondrial cytochrome P-450scc. Mechanism of electron transport by adrenodoxin. The Journal of Biological Chemistry, Apr 1980, s. 3057–61. Dostupné online. DOI: 10.1016/S0021-9258(19)85851-9. PMID 6766943.
- ↑ HASAN, N.; NESTER, E. W.. Purification and properties of chorismate synthase from Bacillus subtilis. The Journal of Biological Chemistry, 1978-07-25, roč. 253, čís. 14, s. 4993–4998. PMID 97285. Dostupné online [cit. 2022-12-28]. ISSN 0021-9258.
- ↑ Prokaryotic assembly factors for the attachment of flavin to complex II. Biochimica et Biophysica Acta (BBA) - Bioenergetics, May 2013, s. 637–47. DOI: 10.1016/j.bbabio.2012.09.003. PMID 22985599.
- ↑ The prokaryotic FAD synthetase family: a potential drug target. Current Pharmaceutical Design, 2013, s. 2637–48. DOI: 10.2174/1381612811319140013. PMID 23116401.
- ↑ a b c d LOV to BLUF: flavoprotein contributions to the optogenetic toolkit. Molecular Plant, May 2012, s. 533–44. DOI: 10.1093/mp/sss020. PMID 22431563.
- ↑ a b c In vivo native fluorescence spectroscopy and nicotinamide adinine dinucleotide/flavin adenine dinucleotide reduction and oxidation states of oral submucous fibrosis for chemopreventive drug monitoring. Journal of Biomedical Optics, 2010, s. 017010–017010–11. DOI: 10.1117/1.3324771. PMID 20210484.
Pozri aj
upraviťZdroj
upraviťTento článok je čiastočný alebo úplný preklad článku Flavin adenine dinucleotide na anglickej Wikipédii.