Sinapse

Sinapses são zonas ativas de contato entre uma terminação nervosa e outros neurônios, células musculares ou células glandulares. Do ponto de vista anatômico e funcional, uma sinapse é composta por três grandes compartimentos: membrana da célula pré-sináptica, fenda sináptica e membrana pós-sináptica. Os principais tipos de contato sináptico são: axo-somático (entre um axônio e o corpo celular), axo-dendrítico (entre um axônio e um dendrito), neuroefetor (entre a terminação nervosa e a célula efetora, fibra muscular lisa, fibra muscular cardíaca ou célula glandular), neuromuscular (entre a terminação nervosa e a fibra muscular esquelética).[1]
Os neurônios nas sinapses
As células nervosas diferenciam-se das demais células do organismo por apresentarem duas propriedades especiais:
- 1) são capazes de conduzir sinais bioelétricos por longas distâncias sem que haja enfraquecimento do impulso ao longo de seu percurso;
- 2) por apresentarem conexões com células musculares (lisas e estriadas), glandulares e outras células nervosas. Estas conexões permitem que sejam produzidas respostas nos músculos cardíaco, liso e esquelético, glândulas exócrinas e neurônios pós-sinápticos através da liberação de substâncias químicas específicas denominadas de neurotransmissores.[2][3]
Os neurônios dispõe-se em cadeias celulares de transmissão e processamento de informações. Um neurônio é composto por três partes: o corpo celular ou soma que é o “centro integrador” dos neurônios um axônio que conduz informações de saídas; e dendritos, que são numerosas projeções ramificadas do corpo celular, recebem e transmitem impulsos nervoso. A forma, o número, e comprimento dos axônios e dendritos e o número de terminais pré-sinápticos (que podem variar de alguns poucos até em torno de 200.000) variam de um neurônio para outro. Estas diferenças fazem os neurônios de diferentes partes do sistema nervoso reagirem de maneira diversa a sinais sinápticos aferentes e executarem muitas funções distintas.[2][3]
Cada sinapse tem duas partes: o terminal axônico da célula pré-sináptica e a membrana da célula pós-sináptica. Em um reflexo neuronal, a informação se transfere da célula pré-sináptica para a célula pós-sináptica. As células pós-sinápticas podem ser neurônios ou não. Na maioria das sinapses entre neurônios, os terminais axônios pré-sinápticos estão próximos dos dendritos ou do corpo celular do neurônio pós-sináptico.[3]
Tipos de sinapse
As sinapses podem ser químicas ou elétricas, em função do tipo de sinal que passará pela célula pré-sináptica e pós-sináptica. As sinapses químicas utilizam mediadores químicos, os neurotransmissores, que medeiam o sinal químico de uma célula pré-sináptica passando pela fenda para uma célula pós-sináptica. As sinapses químicas são as mais utilizadas na transmissão de sinal no sistema nervoso central da espécie humana.[2][3]
As sinapses elétricas transmitem informação instantaneamente de uma célula para outra, com transferência direta de corrente elétrica entre a célula pré-sináptica e a pós-sináptica. Elas são particularmente úteis quando a velocidade e a precisão na transmissão do impulso são fundamentais, como, por exemplo, no músculo cardíaco e no músculo liso.[2][4]
Dentre as sinapses químicas, é possível classificarmos ainda 2 tipos morfológicos ao microscópio eletrônico.[5]
- Tipo I - Apresentam vesículas de forma predominantemente arredondada, com alta densidade pós-sináptica e, segundo a visão clássica, são excitatórias, carregando o neurotransmissor glutamato. Podem ser chamadas também de sinapses assimétricas.
- Tipo II - Apresentam vesículas de forma mais alongada, com menos densidade pós sináptica. Também segundo a visão clássica, são inibitórias, carregando o neurotransmissor GABA. De forma lógica, são chamadas também de sinapses simétricas.
Publicações mais recentes vêm fornecendo novas visões em relação a função dos tipos morfológicos de sinapse, levando em consideração os tipos de receptores pós-sinápticos e o possível papel de sinapses simétricas como locais de comunicação transitória.[6] .
Neurotransmissores
Os neurotransmissores que possuem moléculas pequenas e de ação rápida são aqueles que induzem a resposta mais aguda ao sistema nervoso, como a transmissão de sinais sensoriais para o encéfalo. Na maioria dos casos eles são sintetizados no citosol do terminal pré-sináptico. Alguns exemplos desses neurotransmissores são aminas (serotonina, dopamina), aminoácidos (glicina, glutamato), acetilcolina, gasotransmissores (óxido nítrico, sulfeto de hidrogênio, monóxido de carbono), dentre outros.[2]
Os neuropeptídios provocam ações mais prolongadas, como mudanças em longo prazo no numero de receptores neuronais e provavelmente mudanças em longo prazo no número ou tamanho das sinapses. Eles não são sintetizados no citosol dos terminais pré-sinápticos, mas como partes integrais de grandes moléculas proteicas nos ribossomas, localizados no corpo celular dos neurônios.[2]
A formação de um neuropeptídio é bem mais complexa do que de um neurotransmissor; por isso, é liberado com menor probabilidade do que de um neurotransmissor de pequena molécula. No entanto, a potência de um neuropeptídio é cerca de mil vezes maior do que um neurotransmissor de pequena molécula.[2]
Terminal pré-sináptico: Liberação dos neurotransmissores para a fenda sináptica
O terminal pré-sináptico é separado do terminal pós-sináptico pela fenda sináptica. O terminal pré-sináptico possui dois tipos de estruturas internas para a função excitatória ou inibitória da sinapse: as vesículas transmissoras, que contém os neurotransmissores, e a mitocôndria, que fornece energia necessária para sintetizar novas moléculas de neurotransmissores.[2]
Quando a despolarização de um potencial de ação alcança o terminal pré-sináptico, a mudança no potencial de membrana dá inicio a uma sequência de eventos. A membrana do terminal axônico (terminal pré-sináptico) possui canais de cálcio controlados por voltagem que se abrem em resposta à despolarização da membrana. Como os íons cálcio são mais concentrados no líquido extracelular do que no citosol, eles se movem para dentro da célula. Quando os íons cálcio entram no terminal pré-sináptico, acredita-se que se liguem a proteínas especiais (proteínas do complexo SNARE) presentes em um sítio de liberação na superfície interna da membrana.
Esta ligação, por sua vez, provoca a fusão das vesículas na membrana dos sítios de liberação, permitindo que algumas vesículas contendo os neurotransmissores liberem seu conteúdo na fenda sináptica após cada potencial de ação. As moléculas do neurotransmissor se difundem através da fenda para se ligarem com receptores da célula pós-sináptica. Quando os neurotransmissores se ligam aos seus receptores, uma resposta é iniciada na célula pós-sináptica.[2][3]
Ação dos neurotransmissores no terminal pós-sináptico
-
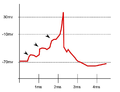 Iniciação de um potencial de ação em um neurônio
Iniciação de um potencial de ação em um neurônio

Os neurotransmissores são responsáveis pelos efeitos eletrofisiológicos excitatórios e inibitórios nas sinapses. Esses neurotransmissores ligam-se a um grande número de proteínas receptoras presentes na membrana do neurônio pós-sináptico, sendo que estas são formadas por combinação de várias subunidades – isso implica a provável existência de dezenas de receptores funcionalmente distintos para alguns neurotransmissores As moléculas desses receptores possuem dois componentes importantes: um componente de ligação que se exterioriza, a partir da membrana, na fenda sináptica, e um componente ionóforo que atravessa toda a membrana pós-sináptica.
Esse componente ionóforo é subdividido em dois tipos: um canal iônico que permite a passagem de íons através da membrana, e um ativador de “segundo mensageiro”, que se projeta para o citoplasma da célula, e tem função de ativar substâncias localizadas no interior do neurônio pós-sináptico.[2][3]
Estes canais iônicos são de dois tipos: canais catiônicos, que permitem a passagem de íons sódio na maioria das vezes, e de íons potássio e/ou cálcio, em menores quantidades; e canais aniônicos, que permitem a passagem de íons cloreto e também de outros íons, em pequenas quantidades. Quando os canais catiônicos se abrem e ocorre a passagem de íons sódio, sua carga positiva provoca a excitação do neurônio, e quando ocorre à passagem de íons de cargas negativas com a abertura dos canais aniônicos, provoca a inibição do neurônio. As substâncias transmissoras provocam a abertura e fechamento desses canais – esse processo de abertura e fechamento ocorre em milésimos de segundos -, fornecendo meios para um controle muito rápido dos neurônios pós-sinápticos.[2][3]
O sistema químico dos receptores ativadores de “segundo mensageiro” possibilita excitação e inibição de efeito prolongado, sendo usado em funções como a memória, por exemplo, que requerem mudanças prolongadas nos neurônios. Entre os diversos tipos de sistemas de segundos mensageiros, os mais comuns são um grupo de proteínas chamadas de proteínas G. Elas estão ligadas a um receptor, e possuem três componentes: componente α, que é a porção ativadora da proteína, e os componentes β e γ, que estão ligados ao componente α. O componente α se desprende dos componentes β e γ durante a ativação por impulso nervoso, ficando livre para se mover no citoplasma da célula, podendo executar várias funções que variam de acordo com a característica específica de cada neurônio.
Essas quatro funções são:
- 1) abertura de canais iônicos específicos na membrana da célula pós-sináptica – promove uma abertura por tempo prolongado do canal, ao contrário dos canais iônicos ativados diretamente;
- 2) ativação do monofosfato de adenosina cíclico (AMPc) ou monofosfato de guanosina cíclico (GMPc) na célula neuronal – AMPc e GMPc responsáveis por ativar maquinaria metabólica altamente específica do neurônio;
- 3) ativação de uma ou mais enzimas intracelulares – estas enzimas podem induzir várias funções químicas específicas das células; (4) ativação da transcrição gênica – provoca a formação de novas proteínas dentro do neurônio.[2]
Receptores inibitórios e excitatórios na membrana pós-sináptica
Um neurotransmissor pode produzir uma transmissão excitatória, que cria um impulso nervoso, ou uma transmissão inibitória, que impede os impulsos nervosos subsequentes. Estes neurotransmissores irão se ligar a receptores determinados na membrana pós-sináptica que quando ativados irão induzir essa excitação ou inibição.[7]
Para a excitação do neurônio é necessário que o potencial interno da membrana pós-sináptica fique mais positivo em relação ao potencial de repouso, o que tem um caráter excitatório. Para tal, pode ocorrer a abertura dos canais de sódio, permitindo o fluxo de um grande número de cargas positivas para o interior do terminal pós-sináptico. Pode também ocorrer uma diminuição da difusão de íons cloreto carregados negativamente para dentro do terminal pós-sináptico ou redução da difusão de íons potássio carregados positivamente para fora do terminal pós-sináptico. Nos dois casos o potencial interno da membrana fica mais positivo. Além disso, mudanças no metabolismo interno do neurônio, no número de receptores ou em propriedades intrínsecas da membrana podem alterar a excitabilidade do neurônio.[2]
Neuromoduladores
São substancias químicas liberadas nas fendas sinópticas que geram efeitos mais lentos e mais a longo prazo do que os efeitos dos neurotransmissores. As ações dos neuromoduladores influenciam o efeito geral dos neurotransmissores, modulando-os. Alguns neurotransmissores, como a noradrenalina, podem agir como neuromoduladores em casos específicos, devido às características de seus efeitos.[8]
Referências
- ↑ Marcus Lira Brandão (2008). As bases biológicas do comportamento. Introdução à neurociência. São Paulo: E.P.U. 244 páginas. ISBN 8512406305
- ↑ a b c d e f g h i j k l m A. C. Guyton, J. E. Hall (2006). Tratado de fisiologia médica 11ª ed. Rio de Janeiro: Elsevier. 1115 páginas. ISBN 8535216413
- ↑ a b c d e f g Dee Unglaub Silverthorne (2010). Fisiologia humana. Uma abordagem integrativa 5ª ed. Porto Alegre: Artmed. 992 páginas. ISBN 9788536322841
- ↑ Margarida de Mello Aires (2008). Fisiologia 3ª ed. Rio de Janeiro: Guanabara-Koogan. 1252 páginas. ISBN 9788527713689
- ↑ Gray EG (1959) Axo-somatic and axo-dendritic synapses of the cerebral cortex: an electron microscope study. J. Anat. (Lond.), 93:420-433.
- ↑ Klemann CJ1, Roubos EW (2011) The gray area between synapse structure and function-Gray's synapse types I and II revisited Synapse, 65(11):1222-30.
- ↑ Gerard Jerry Tortora, Bryan Derrickson (2010). Princípios de anatomia e fisiologia 12ª ed. Porto Alegre: Artmed. 1256 páginas. ISBN 8527716534
- ↑ Lent, Roberto. «4 - Chips Neurais». Cem Bilhões de Neurônios 2a ed. [S.l.: s.n.]
Ver também