

Apparato visivo

L'apparato visivo dei vertebrati è formato da due organi esterni pari e simmetrici posti nella regione anteriore della testa, gli occhi, considerati come un'appendice dell'encefalo, sia per derivazione embriologica, sia per una serie di correlazioni funzionali, come la capacità integrativa propria delle strutture nervose, che si ritrova a livello della retina. L'apparato visivo nel suo insieme è un sistema di elaborazione specializzato formato da un insieme di zone cerebrali diverse[1].
Considerazioni preliminari
[modifica | modifica wikitesto]Alla fine del 1600 William Molyneux pose la questione: se una persona nata cieca avesse potuto riacquistare la vista avrebbe potuto riconoscere gli oggetti con il nuovo senso? Oggi sappiamo che adeguate tecniche operatorie che possono risolvere i problemi oculari, se eseguite a molti anni dalla nascita, danno risultati deludenti in quanto le aree cerebrali non hanno sviluppato un'educazione visiva.
Ciò che ogni essere vivente vede è il frutto dell'elaborazione centrale di informazioni provenienti dalle retine a confronto con le esperienze precedenti. Queste vengono "pesate e smistate" dal lobo occipitale (in particolare dalle aree di Brodmann nn. 17, 18 e 19), arricchite di significato e interfacciate con le altre informazioni dalle aree associative. Si avrà infine un'immagine tridimensionale, che corrisponde a suoni, sensazioni ed emozioni.
Tutto ciò che “noi vediamo” è il frutto della luce riflessa dagli oggetti, passata attraverso le pupille, distorta e ribaltata dai mezzi diottrici oculari e proiettata sulla superficie curva bidimensionale della retina, che si comporta come un trasduttore tra l'energia dei fotoni e il segnale trasmesso tramite potenziali d'azione. La retina, inoltre, è in grado di percepire colori e di dare informazioni precise e accurate riguardo allo spazio che ci circonda solo nella fovea. Inoltre, le retine di entrambi gli occhi hanno un punto cieco, dove l'immagine non può essere percepita. Tuttavia, non percepiamo un'immagine frammentata, distorta o bidimensionale: poiché è necessario un processo integrativo ed elaborativo che gestisce le innumerevoli informazioni provenienti dalla retina. Questi processi sono svolti dal cervello, il vero e proprio organo della visione[2].
Per comprendere come il cervello operi nei processi visivi, basta soltanto considerare le cosiddette illusioni ottiche quali il triangolo di Kanizsa, le figure di Muller-Lyer o di Edgar Rubin. Queste illusioni mettono in luce il ruolo centrale del cervello come centro della visione: costruisce l'immagine secondo i propri canoni, sfrutta oggetti nel campo visivo per ricavare informazioni su altri (si pensi alla prospettiva; la visione su una stessa immagine di due persone, una più piccola e una più grande, fa concludere che quella più grande si trovi più vicina a noi e viceversa), analizza le profondità prendendo per assunto che la luce provenga dall'alto, così da far percepire una semisfera più chiara in basso che in alto come una semisfera concava e nel caso contrario convessa.
Il cervello costruisce quindi l'immagine visiva con un processo attivo, e non passivo come ritenuto per millenni, in accordo con le moderne teorie sulla percezione e della Psicologia della Gestalt.
Anatomia e Fisiologia
[modifica | modifica wikitesto]L'apparato visivo è formato da:
- Occhio ed annessi;
- Nervo ottico;
- Tratto ottico;
- Pretetto, Ganglio ciliare, Nucleo oculomotore accessorio (di Edinger-Westphal), vie ortosimpatiche e parasimpatiche;
- Collicolo superiore;
- Corpo genicolato laterale;
- Radiazioni ottiche o calcarine;
- Corteccia visiva, che si divide in primaria, secondaria e terziaria;
- Corteccia associativa, teorizzata da Paul Flechsig nel 1907.
Verranno in seguito analizzati in termini fisiologici tutti gli organi elencati.
Occhio
[modifica | modifica wikitesto]Coni e bastoncelli
[modifica | modifica wikitesto]| Neurotrasmettitori e Neuropeptidi Retinici | |
|---|---|
| Neurotrasmettitori | |
| Glutammato | Principale neurotrasmettitore retinico, utilizzato dalla maggior parte delle cellule. |
| GABA | Secreto da pochi tipi di cellule orizzontali ed amacrine. |
| Glicina | Secreto da cellule orizzontali ed amacrine; anche bipolari. |
| Dopamina | Secreto da cellule amacrine A18 |
| Acetilcolina | Secreto da alcune cellule amacrine. Le cellule gangliari Y hanno recettori muscarinici e recettori nicotinici. |
| Neuropeptidi | |
| Sostanza P | Si trova in un tipo di cellule amacrine e nelle cellule gangliari. |
| Somatostatina | Si trova in alcune cellule amacrine e in neuroni di associazione di Cajal. |
| CRH | È contenuto in cellule amacrine, probabilmente le stesse che secernono anche somatostatina. |
| VIP | Il peptide vaso attivo intestinale è prodotto da una popolazione di cellule amacrine A12. |
| Neuropeptide Y | Si ritrova in alcune cellule Amacrine responsabili dei circuiti ON e OFF.[3] |
Per la struttura microscopica della retina si rimanda all'anatomia; in questa sede verranno presi in considerazione coni, bastoncelli, cellule amacrine, cellule orizzontali, cellule bipolari e cellule gangliari e i rapporti reciproci necessari alla comprensione dei meccanismi di modulazione e trasmissione del segnale luminoso.
Qui a fianco sono riportate le diverse sostanze che trasducono le informazioni che si generano nei coni e nei bastoncelli. Questi ultimi hanno il compito di "tradurre" il segnale luminoso in segnale elettrico e sono, dopo l'epitelio pigmentato (formato da melanociti ricchi di pigmento scuro in grado di fare da camera oscura), le cellule più esterne (più lontano dal centro) dell'occhio. Inoltre, di tutto il recettore, solo l'articolo esterno (segmento cellulare più esterno) è in grado di recepire l'energia luminosa. Di seguito sono riportate delle nozioni utili da tener presente per la comprensione dei fenomeni descritti successivamente.
Nell'articolo esterno dei bastoncelli sono presenti numerosi dischi membranosi che galleggiano nel citoplasma, dove viene espressa una proteina di membrana a 7 segmenti, l'opsina. L'opsina contiene il retinale; il complesso opsina-retinale viene detto rodopsina. La stessa cosa nei coni, dove però i dischi membranosi non sono galleggianti, ma sono dipendenze della stessa membrana cellulare. Al buio, il retinale della rodopsina è in conformazione 11-cis; se colpito da un fotone, può diventare retinale tutto-trans. Questo composto è in grado di cambiare la conformazione sterica della opsina, che così attivata, può attivare molte proteine G dette trasducine, le quali sono in grado di attivare molte GMP-ciclico fosfodiesterasi, enzimi che idrolizzano in 5'-GMP un'enorme quantità di GMP-ciclico (cGMP). Dunque, se c'è luce, la concentrazione plasmatica di cGMP diminuisce criticamente.
Nell'articolo esterno sono presenti numerosi canali per il sodio e per il calcio. Se questi canali sono aperti nel citoplasma ci sarà una notevole concentrazione di questi soluti, mantenuta tuttavia adeguata dalla pompa sodio-potassio e da pompe per il calcio che lo estrudono costitutivamente. Se i canali del sodio/calcio sono aperti, allora la conduttanza specifica per questi ioni è alta: questo è il motivo per cui in un cono o bastoncello il potenziale di riposo di membrana (di riposo, ovvero al buio, non stimolato) è più depolarizzato che in un normale neurone (-40mV). Al buio, dunque, la cellula sarà ricca di sodio e di calcio; quest'ultimo ione è il principale responsabile dell'esocitosi delle vescicole contenenti glutammato. È importante inoltre sottolineare che questi canali per il sodio/calcio, al buio, sono mantenuti aperti dal legame con il GMP ciclico che si lega nel lato citosolico del canale e che sia al buio che alla luce sono presenti nell'articolo interno un gran numero di canali per il potassio sempre aperti, che garantiscono una corrente uscente di potassio costante.
Dunque, se arriva luce nei coni e nei bastoncelli allora ci sarà molto meno cGMP nella cellula. Tanto meno cGMP ci sarà nella cellula tanti meno canali per il sodio/calcio saranno aperti. In presenza di luce, pertanto, i canali per il sodio/calcio tenderanno a chiudersi. La chiusura dei canali per il sodio/calcio comporta una serie di conseguenze innescate dal crollo della conduttanza specifica per questi ioni. Tuttavia la conduttanza del potassio rimarrà costante e garantirà una corrente uscente continua. Questo porta ad una iperpolarizzazione del recettore, il quale passerà da -40 mV a -75 mV, potenziale al quale il recettore non secerne neurotrasmettitore.
In conclusione, i recettori della retina sono cellule del tutto particolari che, a differenza della maggior parte dei neuroni, trasmettono il segnale come uno stimolo elettrotonico iperpolarizzante invece che depolarizzante. Dunque, se si ha iperpolarizzazione, si ha segnale.
Adattamento
[modifica | modifica wikitesto]Se si passa da una stanza molto buia, ad una stanza molto illuminata, si ha il fenomeno dell'abbagliamento, che di solito dura pochi secondi. Tuttavia, è stato spiegato che uno stimolo luminoso provoca una iperpolarizzazione. È logico pensare che, passando da una stanza buia ad una stanza molto illuminata, ci sarà una iperpolarizzazione massiva di tutti i fotorecettori, che a questo punto non potranno più iperpolarizzarsi ulteriormente e trasmettere, ad esempio, i particolari della stanza, cosa notoriamente non vera. Infatti, mentre siamo abbagliati, hanno luogo una serie di eventi che ci permettono di distinguere ulteriori particolari. In primo luogo, un riflesso parasimpatico, successivamente descritto, fa restringere la pupilla, ostacolando l'eccessiva entrata di luce.
In secondo luogo, si ha un fenomeno di adattamento cellulare, dato che:
- in un recettore, la guanilato ciclasi, che catalizza la formazione di cGMP, è inibita dal calcio;
- in un recettore, la cGMP-fosfodiesterasi, che catalizza la formazione di GMP, è favorita dal calcio;
- in un recettore colpito da stimolo luminoso, si chiudono i canali del sodio/calcio. Questi minerali tuttavia vengono estrusi attivamente dal citosol per opera di pompe specifiche. Quindi, la stimolazione luminosa fa abbassare la concentrazione citosolica di sodio e calcio.
Dunque, date queste premesse, se un recettore è illuminato, la concentrazione di calcio diminuisce. Se la concentrazione di calcio diminuisce, la concentrazione di cGMP aumenta, poiché si riattiva la guanilato ciclasi e si inibisce la cGMP-fosfodiesterasi. Se la concentrazione di cGMP aumenta, allora si riaprono i canali per il sodio/calcio, depolarizzando la cellula. Questa depolarizzazione verso il potenziale di riposo di -40 mV, rende possibile una nuova stimolazione iperpolarizzante e dunque i recettori possono ritornare a segnalare, ad esempio, i dettagli di una stanza molto illuminata.
Cellule gangliari e la rete degli interneuroni
[modifica | modifica wikitesto]L'iperpolarizzazione di coni e bastoncelli, dovuta allo stimolo luminoso, porta alla cessazione della secrezione di glutammato, per il quale cellule più interne e in sinapsi con gli stessi coni o bastoncelli, le cellule bipolari, hanno recettori di membrana.
Nelle cellule bipolari, non si ha ancora potenziale d'azione e l'impulso elettrico, iperpolarizzante o depolarizzante, generato dai canali sensibili al glutammato, viene condotto elettrotonicamente.
Se una cellula bipolare viene depolarizzata, essa secernerà neurotrasmettitori eccitatori alle cellule gangliari, poste più internamente, nelle quali si genererà il potenziale d'azione.

Cellule gangliari
[modifica | modifica wikitesto]Le cellule gangliari sono le prime cellule del sistema visivo nelle quali si generano potenziali d'azione. I loro assoni proiettano al pretetto, per le risposte vegetative, al collicolo superiore, affinché le informazioni visive vengano interfacciate con le altre informazioni sensoriali e al corpo genicolato laterale del talamo, per entrare in sinapsi con neuroni che proiettano alla corteccia striata. Questi assoni convergono tutti verso il punto cieco della retina, là dove si uniscono per formare il nervo ottico.
Le cellule gangliari possono essere distinte in cellule M (per magnae o grandi) e cellule P (per parvae o piccole). Le cellule M sono connesse con un gran numero di coni e bastoncelli e per questo sono in grado di fornire informazioni sul movimento di un oggetto o su rapide variazioni del contesto luminoso. Le cellule P sono connesse con un minor numero di recettori e sono adatte a fornire informazioni sulla forma e sul colore di un oggetto.
Questa distinzione anatomo/funzionale è importantissima: gli assoni provenienti dalle cellule M e dalle cellule P rimangono segregati anche a livello centrale. Questi infatti proiettano a zone distinte del corpo genicolato laterale: inoltre, dal corpo genicolato laterale, le informazioni provenienti dai 2 tipi cellulari, vengono smistate a zone diverse della corteccia striata. Da qui, le informazioni prendono due vie distinte; la via ventrale, per le informazioni in gran parte provenienti dalle cellule P e la via dorsale, per le informazioni in gran parte provenienti dalle cellule M.
Campi recettivi: cellule centro-on e cellule centro-off
[modifica | modifica wikitesto]Sia le cellule P che le cellule M possono essere distinte ulteriormente in cellule gangliari centro-on e cellule gangliari centro-off. Al fine di comprendere la distinzione occorre definire il concetto di campo recettivo.
Per un neurone gangliare delle radici posteriori del midollo spinale (le cellule deputate alla percezione degli stimoli tattili-termici-dolorifici) il campo recettivo è quella porzione di cute che se stimolata opportunamente produce la massima attivazione dello stesso neurone gangliare. Analogamente, per una cellula gangliare retinica il campo recettivo è quella porzione di retina che se stimolata produce la massima risposta della stessa cellula gangliare. Analogamente ai campi recettivi presenti nella cute, i campi recettivi retinici possono essere più o meno estesi e a seconda della loro grandezza, forniscono informazioni più o meno precise sulla natura di uno stimolo.
In particolare, più sono piccoli i campi recettivi, maggiore sarà la loro densità e migliore sarà la discriminazione di un dato oggetto (per la cute, ad esempio, i polpastrelli hanno campi recettivi minuscoli e hanno dunque un'enorme possibilità discriminativa). Si comprende ora che i campi recettivi delle cellule M sono molto grandi, mentre quelli delle cellule P sono piccoli ed è questo il motivo della loro elevatissima capacità discriminativa. Ad una cellula gangliare sono collegati un certo numero di recettori; maggiore sarà il numero di questi recettori, maggiore sarà il campo recettivo.
In particolare, si ha che un certo numero di recettori in un'area circolare di retina è connesso con un certo numero, minore, di cellule bipolari, le quali sono connesse con una sola cellula gangliare.
A questo punto è lecito aspettarsi che la stimolazione luminosa di questa area circolare di retina (il campo recettivo), produrrà una certa risposta della cellula gangliare. La risposta della cellula gangliare allo stimolo luminoso varia a seconda del tipo di cellula che viene investita dallo stimolo luminoso
Infatti, se viene investito il campo recettivo di una cellula centro-on dallo stesso stimolo luminoso che investe il campo recettivo di una cellula centro-off, si produce una risposta diversa.
Nella figura sono illustrate le differenze di risposta di queste cellule per uno stesso stimolo luminoso. Si comprende quindi che:
- Le cellule centro-on rispondono massivamente (nel senso di numero di potenziali d'azione per secondo) per uno stimolo luminoso localizzato nel centro del loro campo recettivo, mentre le cellule centro-off rispondono massivamente per uno stimolo luminoso localizzato alla periferia del loro campo recettivo.
- Condizioni opposte al punto 1, provocano l'annullamento dei potenziali d'azione in entrambe le cellule.
- Per ogni cellula il centro dà una risposta contrastante alla periferia. Questo comporta che un'illuminazione diffusa produce solo una debole risposta in ambedue le cellule.
Queste proprietà di cellule centro-on e cellule centro-off permettono 2 cose:
- Una adeguata e precisissima percezione dei contrasti: l'alternarsi e il sovrapporsi dei campi recettivi di queste stesse cellule, produce zone ad elevatissima discriminazione luminosa. In una stessa piccola zona di un millimetro quadrato, possono esserci tantissimi campi recettivi di entrambe le cellule. Un quadrato a linee tratteggiate ci appare diverso da un quadrato a linee punteggiate proprio perché punti e linee scure sono zone che riflettono meno la luce e che quindi possono essere percepite in maniera definitissima dall'alternarsi di campi recettivi centro-on e centro-off. Questo fenomeno è di fondamentale importanza poiché il sistema visivo costruisce l'immagine a partire dai contorni degli oggetti (dovuti al contrasto dell'oggetto con il mondo che lo circonda), deducendo in seguito movimento, profondità e poi colore.
- Una adeguata percezione dei cambiamenti di illuminazione. Dall'immagine infatti si deduce che le cellule centro-on riescano a percepire molto bene i rapidi aumenti di illuminazione; al contrario le cellule centro-off percepiscono al meglio le rapide diminuzioni di illuminazione.

Interneuroni
[modifica | modifica wikitesto]In precedenza sono stati descritti i fenomeni associati alla presenza di campi recettivi retinici centro on e centro off. È stato chiarito come l'integrazione delle informazioni derivanti dalle cellule gangliari competenti per questi campi sia importante per la discriminazione dei contrasti, fondamentale per l'analisi visiva. Tuttavia, non sono né le cellule gangliari né i recettori i responsabili di questa discriminazione: le cellule responsabili del fenomeno sono gli interneuroni della retina, che tramite una rete di contatti reciproci, sono in grado di costituire “l'impalcatura” necessaria alla costruzione dei campi recettivi centro on e centro off.
Prima di procedere con un'analisi più approfondita dei fenomeni di fisiologia molecolare occorre ricordare che esistono diversi specie di interneuroni nella retina, tra cui le cellule orizzontali, le cellule amacrine e le cellule bipolari. Si rimanda all'anatomia microscopica della retina per la comprensione dei rapporti specifici che questi interneuroni contraggono tra loro, con i recettori e con le cellule gangliari.
Tuttavia uno schema anatomico generale può facilitare la comprensione, prendendo in analisi una retina si ha che:
- I recettori (coni e bastoncelli) sono le cellule più esterne (se si escludono i melanociti dell'epitelio pigmentato);
- I recettori contraggono sinapsi con delle cellule più interne, le cellule bipolari;
- Le cellule bipolari scaricano il segnale elettrotonico stimolato dai recettori alle cellule gangliari. Se scaricano il segnale a cellule gangliari centro-on, la cellula bipolare si definisce cellula bipolare centro-on. Se scaricano il segnale a cellule gangliari centro-off, la cellula bipolare si definisce cellula bipolare centro-off;
- Le cellule bipolari centro-on hanno canali per il potassio (iperpolarizzanti) aperti tramite il legame con il glutammato. Le cellule bipolari centro-off hanno canali per il sodio (depolarizzanti) aperti tramite il legame con il glutammato.
- Le cellule bipolari dunque fanno da ponte tra i recettori (più esternamente) e le cellule gangliari (più internamente);
- Gli assoni delle cellule gangliari escono dalla retina formando il nervo ottico;
- I recettori sono collegati tra di loro mediante le cellule orizzontali.
Inoltre è opportuno ricordare che, tra tutti i neuroni, solo i neuroni depolarizzati sono in grado di aprire i canali per il calcio e solo il calcio è in grado di scatenare la fusione di vescicole secretorie, pertanto il neurotrasmettitore può essere secreto solo da neuroni in stato depolarizzato.
Si noti anche che tra tutte le cellule della retina, solo le cellule gangliari, siano esse P o M, centro on o centro off, sono in grado di produrre potenziali d'azione. Tutte le altre sono cellule che in risposta a stimoli conducono elettrotonicamente variazioni del potenziale di membrana.
La figura qui a fianco fornisce 2 esempi tipici di facile comprensione, illustranti la via diretta e la via indiretta. Nel primo caso si analizza un recettore del centro del campo recettivo, nel secondo un recettore della periferia del campo recettivo.
Via diretta
[modifica | modifica wikitesto]Si prenda un bastoncello del centro;
Se colpito dall'onda luminosa, esso (si veda sopra) tende alla iperpolarizzazione. Questo è uno stato che impedisce il rilascio di neurotrasmettitore; l'assenza di neurotrasmettitore ha un duplice effetto:
- Senza glutammato i canali ionici depolarizzanti delle cellule bipolari centro off si chiudono; la cellula ne risulterà iperpolarizzata (non secernerà neurotrasmettitore alle cellule gangliari centro off).
- Senza glutammato i canali metabotropici iperpolarizzanti delle cellule bipolari centro on si chiudono; la cellula ne risulterà depolarizzata (infatti se si chiudono i canali per il potassio la cellula viene “tirata” verso il potenziale di equilibrio del sodio) e secernente neurotrasmettitore alle cellule gangliari centro-on.
Analizzando gli effetti, si avrà che, in presenza di luce centrale, gli assoni provenienti dalle cellule gangliari centro on condurranno treni di potenziali d'azione, informando strutture superiori della presenza di luce nel centro; inoltre gli assoni provenienti dalle cellule gangliari centro off avranno una scarica molto minore, informando i centri superiori della presenza di luce nel centro (una sorta di rinforzo e conferma reciproca).
Se invece il bastoncello non è colpito dall'onda luminosa si trova in uno stato di riposo, “costitutivamente” depolarizzato, secernente “costitutivamente” un neurotrasmettitore, il glutammato.
Tale neurotrasmettitore secreto dai bastoncelli ha un duplice effetto sulle cellule bipolari:
- Apre i canali ionici depolarizzanti nelle cellule bipolari centro off; queste si depolarizzeranno secernendo neurotrasmettitore depolarizzante alle cellule gangliari centro-off.
- Apre i canali iperpolarizzanti nelle cellule bipolari centro-on; queste tenderanno alla iperpolarizzazione e dunque non secerneranno neurotrasmettitore depolarizzante alle cellule gangliari centro-on.
Analizzando gli effetti anche in questo caso, si avrà che, in assenza di luce centrale, gli assoni provenienti dalle cellule gangliari centro-off condurranno treni di potenziali d'azione, informando strutture superiori dell'assenza di luce nel centro; inoltre gli assoni provenienti dalle cellule gangliari centro-on avranno una scarica molto minore, informando i centri superiori dell'assenza di luce nel centro (anche qui si ha dunque rinforzo e conferma).
Questi primi due fenomeni fanno riferimento alla parte superiore della figura qui a fianco.
Via Indiretta
[modifica | modifica wikitesto]La parte inferiore può risultare di difficile comprensione se non viene “letta” con le seguenti nozioni:
- Le cellule orizzontali ed amacrine sono interneuroni inibitori, ovvero tendono ad iperpolarizzare post-sinapticamente le cellule bersaglio;
- Le cellule bersaglio di questi interneuroni sono i recettori (coni e bastoncelli);
- Questi interneuroni connettono i recettori del campo recettivo periferico con i recettori del campo recettivo centrale.
Si prenda un campo recettivo centro-off, stimolato adeguatamente da un fascio di luce periferico; si avrà che:
- Il bastoncello periferico viene iperpolarizzato;
- Il bastoncello periferico trasmette l'iperpolarizzazione ad una cellula orizzontale;
- La cellula orizzontale è iperpolarizzata e dunque inibita ad inibire;
- La cellula postsinaptica (il bastoncello centrale del campo recettivo) alla cellula orizzontale è dunque libera di secernere il neurotrasmettitore (glutammato);
- Questo comporta l'attivazione delle cellule bipolari centro off, secernenti neurotrasmettitore alle cellule gangliari centro-off;
- I treni di potenziali di azione informeranno dunque i centri superiori della presenza di luce nella periferia (si noti che questo segnale è estremamente rinforzato dall'assenza di luce al centro).
Gli stessi fenomeni avvengono in senso opposto nei campi recettivi centro-on.

Retina
[modifica | modifica wikitesto]Per la trattazione “fisiologica” occorre invece intendere la retina come un “epitelio sensitivo” che percepisce e interpreta l'informazione luminosa proveniente dalla pupilla.
Inoltre l'informazione luminosa viene ribaltata e proiettata su una superficie di una semisfera concava con capacità discriminative non uniformi; quello che ne risulta è sicuramente un'immagine distorta.
Questo è un punto importante, che focalizza l'attenzione sul ruolo primario del cervello sulla costruzione di un'immagine coerente, “piana” e dettagliata. La massima capacità discriminativa è localizzata nella fovea, una zona della retina con la massima concentrazione di coni.
La fovea è il punto della retina con maggiore discriminazione spaziale; non solo, è anche il punto in cui si ha la maggiore discriminazione e il maggior confronto cromatico (si veda più avanti la trattazione della visione dei colori). Per sperimentare questo fatto basta chiedere a qualcuno di mostrarci dei colori (cartoncini colorati) mentre siamo impegnati a fissare un qualche cosa lontana dalla persona. Benché la persona con i cartoncini colorati cada nel nostro campo visivo, più questa è lontana dal punto di fuoco più sarà difficile per noi distinguere i colori.
Per ogni occhio, la fovea è disposta lateralmente al punto cieco (vedi figura qui a fianco), zona di uscita degli assoni provenienti dalle cellule gangliari; è dunque logico che questa piccola zona non è in grado di recepire gli stimoli visivi (da cui il nome).
È da notare che il sistema motorio oculare tende a mantenere l'occhio in una posizione tale da consentire soprattutto alla fovea di analizzare il particolare osservato. Si può perciò affermare (è un'evidenza sperimentale) che il sistema visivo muove la fovea verso un particolare e ne costruisce un contesto appropriato intorno.
Si immagini di osservare una figura. Immediatamente le fovee vengono mosse lungo tutti i punti della figura che attirano l'attenzione (in un volto, ad esempio, i punti di interesse sono gli occhi, il naso, la bocca, gli zigomi); in seguito i movimenti diventano sempre più lenti fino a fissare un particolare che ci interessa (punto di fuoco). A questo punto la corteccia comincia a costruire l'immagine visiva, soprattutto tramite la serie di “scatti veloci” impressi in precedenza dai movimenti delle fovee e in parte minore dal contesto analizzato dalle parti periferiche alla fovea.
Il sistema motorio oculare è dunque impegnato costantemente per muovere la fovea. Questi movimenti sono detti saccadici o semplicemente "saccadi", determinati da complessi schemi motori integrati e veicolati da nuclei quali il collicolo superiore in base alle informazioni che provengono dalla corteccia. Si possono raggiungere velocità di movimento dell'ordine di 900 gradi al secondo, accompagnati da soste di fissazione che durano in media 300 ms. È interessante notare che in questi tempi, il sistema visivo della corteccia non è in grado di derivare un quadro completo dell'immagine, ma riceve informazioni "abbozzate" della scena visiva, quelli che, in precedenza, sono stati chiamati, per comprensione, "scatti veloci".
Si potrebbe erroneamente dedurre che le parti periferiche della retina siano poco importanti; al contrario, queste zone, ricche di bastoncelli, sono le maggiori responsabili della percezione del movimento, ma anche dei contrasti (questo soprattutto in visione crepuscolare) e della profondità.
Inoltre per ogni retina è possibile distinguere due parti: una emiretina nasale (ovvero la metà retina di un occhio prospiciente verso il naso) e una emiretina temporale.
La trattazione specifica delle conseguenze fisiologiche di questa divisione avverrà in seguito. Ciò che deve essere invece chiaro a questo livello è che le informazioni visive che cadono sull'emiretina nasale sono quelle che arrivano dal campo visivo laterale; le informazioni visive che cadono sull'emiretina temporale sono quelle che arrivano dal campo visivo mediale.
Ogni emiretina nasale è, per ciascun occhio, competente per la parte di campo di visivo più esterna, mentre l'emiretina temporale è competente per la parte di capo visivo più interna. Queste informazioni prevenienti da ogni singola emiretina rimangono segregate per l'intero tragitto nelle vie visive centrali, così come lo rimangono le informazioni distinte veicolate dagli assoni delle cellule gangliari P e delle cellule gagliari M.

Nervo ottico e tratto ottico
[modifica | modifica wikitesto]Ogni nervo ottico è composto dagli assoni delle cellule gangliari che risiedono nella retina nasale e nella retina temporale di un singolo occhio. Questo comporta che la lesione di un nervo ottico provoca cecità nell'emicampo visivo ipsilaterale alla lesione.
Il tratto ottico è una continuazione del nervo ottico, che decorre dal chiasma ottico (nel quale si ha un parziale incrociamento delle fibre nervose provenienti dalle due emiretine di ciascun occhio) al corpo genicolato laterale del talamo.
Il tratto ottico di destra è composto dalle fibre nervose provenienti dall'emiretina temporale dell'occhio destro e dall'emiretina nasale dell'occhio sinistro; viceversa, il tratto ottico di sinistra è composto dalle fibre provenienti dall'emiretina temporale dell'occhio sinistro e dall'emiretina nasale dell'occhio destro. In pratica, ciascun tratto ottico contiene la rappresentazione completa dell'emicampo visivo controlaterale. Si avrà, dunque, che la corteccia visiva destra "vedrà" il campo visivo di sinistra, mentre la corteccia visiva sinistra "vedrà" il campo visivo di destra.
Una lesione a carico del tratto ottico produce emianopsia omonima controlaterale, cioè la cecità della stessa metà del campo visivo di ciascun occhio.
Le informazioni veicolate dai tratti ottici hanno fondamentalmente 3 destinazioni:
- Pretetto; per le risposte vegetative, riflesso della pupilla e del cristallino.
- Ipotalamo: regolazione dei ritmi circadiani.
- Collicolo superiore; per essere interfacciate con le altre informazioni sensitive, interviene nel controllo dei movimenti della testa e degli occhi.
- Corpo genicolato laterale; per essere proiettate alla corteccia visiva.
Verranno ora prese in considerazione queste importanti stazioni.

Pretetto
[modifica | modifica wikitesto]La zona del pretetto è formata da un gruppo di nuclei posti anteriormente e superiormente al collicolo superiore, nel punto in cui il mesencefalo si continua con il talamo.
Questi nuclei sono importanti poiché sono i principali responsabili delle risposte vegetative connesse con l'apparato visivo; queste risposte sono il riflesso di costrizione e di dilatazione della pupilla, entrambi consensuali, mediati rispettivamente dal sistema parasimpatico e da sistema simpatico.
Sono inoltre importanti i riflessi di puntamento in risposta ad uno stimolo visivo localizzato perifericamente al punto di fissazione foveale; tuttavia questi ultimi hanno sede solo in parte nel pretetto, poiché i movimenti oculari sono in gran parte governati da stimoli provenienti dal collicolo superiore, dalla corteccia e da alcuni nuclei che, in generale, integrano e interfacciano le diverse informazioni sensoriali, evocando una risposta di idoneo movimento oculare (si pensi ad un suono o ad uno stimolo tattile che catturano la nostra attenzione visiva). Di seguito e in figura sono trattate le importanti risposte che hanno sede nel pretetto; le risposte evocate dalla stimolazione collicolare e corticale sono trattate successivamente.
Riflesso di costrizione della pupilla
[modifica | modifica wikitesto]Il riflesso di costrizione della pupilla è un riflesso mediato dal sistema parasimpatico molto importante indagato nella pratica clinica neurologica, utile affinché il soggetto non venga abbagliato prima che entrino in atto tutti i processi di adattamento descritti precedentemente.
Una luce intensa è in grado di provocare questo riflesso innescando un processo che ha come sede principale i nuclei del pretetto. Questi proiettano assoni verso il nucleo di Edinger-Westphal (o nucleo oculomotore accessorio), un piccolo nucleo parasimpatico posto nelle immediate vicinanze del nucleo oculomotore. Il nucleo di Edinger-Westphal proietta assoni pregangliari verso il ganglio ciliare. È importante notare che queste fibre pregangliari "viaggiano insieme" al nervo oculomotore fino al ganglio ciliare. Dal ganglio ciliare si originano i nervi ciliari brevi, fibre parasimpatiche postgangliari che stimolando il muscolo costrittore della pupilla ne provocano la contrazione.
Riflesso di dilatazione della pupilla
[modifica | modifica wikitesto]Il riflesso di dilatazione della pupilla, anch'esso indagato nella pratica neurologica, è un riflesso mediato dal sistema simpatico molto importante, utile nelle condizioni di poca luce ambientale. Permette infatti l'aumento del diametro della pupilla, consentendo alla retina di "lavorare" con un maggiore afflusso di informazioni luminose.
Le poche informazioni luminose che arrivano al pretetto stimolano i neuroni di questi nuclei a contattare, tramite un collaterale della Via Tetto-Spinale laterale, i motoneuroni viscerali della lamina VII presenti a livello di C8-T1 che vanno a costituire il centro cilio-spinale di Budge. Dal centro ciliospinale si dipartono fibre simpatiche pregangliari che risalgono i gangli paravertebrali fino a contattare sinapticamente i neuroni del ganglio cervicale superiore.
Da questi si dipartono fibre simpatiche postgangliari che salgono parallelamente all'arteria carotide interna, fino a raggiungere il plesso carotico interno e da qui il plesso cavernoso a ridosso dell'anello tendineo comune. Parte delle fibre si portano successivamente al ganglio ciliare (radice simpatica del ganglio ciliare) e da qui giungono, senza contrarre sinapsi nel ganglio, al muscolo dilatatore della pupilla, tramite i nervi ciliari brevi (che possono essere dunque sia simpatici che parasimpatici). La restante parte delle fibre, in corrispondenza del plesso cavernoso, entra in anastomosi col nervo naso-ciliare (collaterale del nervo oftalmico), giungendo poi all'iride per mezzo del nervo ciliare lungo.
Riflessi di puntamento
[modifica | modifica wikitesto]Se immagini di stare a guardare un punto di fisso di un paesaggio, le fovee si dispongono affinché la proiezione di quel particolare fissato cada perfettamente su di esse. L'immagine della maggior parte del paesaggio cade dunque in zone retiniche che non hanno le stesse proprietà discriminative della fovea. Tuttavia è importante, per l'uomo e per l'animale, riuscire ad avvertire nel paesaggio "di sfondo" che circonda il fuoco movimenti rapidi e improvvisi che possono comportare pericolo. L'avvertimento della perturbazione è analizzato dalla corteccia, dal collicolo e dal pretetto.
Istintivamente il pretetto contatta i nuclei oculomotori[4] (poco), ma soprattutto i motoneuroni dei muscoli cervicali, sincronizzando e stimolando la loro azione in modo da portare il capo (e dunque occhi e fovee) verso il punto di perturbazione rilevato.
Le vie discendenti che dal pretetto vanno a contattare questi motoneuroni sono la via tetto-spinale mediale (in larga parte) e parte della via tetto-spinale laterale, utilizzata soprattutto dal riflesso di dilatazione delle pupille.
Detto questo, occorre considerare che sebbene si tratti di un vero e proprio riflesso istintivo, questo può essere soverchiato dalla corteccia (dalla volontà), nel momento in cui il soggetto decide di non rispondere ad uno stimolo che invade il suo campo visivo. Questo non succede invece con il riflessi di costrizione e dilatazione della pupilla, assolutamente non allacciati al controllo volontario. Cosa importante è ricordare che i riflessi di puntamento utilizzano preferenzialmente i muscoli del collo, poiché potenti e in grado di resistere alla fatica. Non è di secondaria importanza il fatto che, ruotando il capo invece degli occhi, il soggetto si ritrova anche orecchi, naso e bocca indirizzati verso lo stimolo, elementi che possono aiutare a comprendere la natura della perturbazione.

Rivalità binoculare
[modifica | modifica wikitesto]La rivalità binoculare è la situazione che si viene a creare quando ad un occhio viene presentata contemporaneamente un'immagine diversa dall'altro occhio, tanto che la nostra percezione passa da una all'altra immagine per volta[5].
Collicolo superiore
[modifica | modifica wikitesto]Nell'immagine qui a fianco è rappresentata la superficie posteriore del tronco dell'encefalo dopo asportazione del cervelletto.
In blu è possibile apprezzare superiormente il corpo genicolato laterale e inferiormente il collicolo superiore; quest'ultimo è un'importante stazione mesencefalica corrispondente al tetto ottico dei vertebrati inferiori.
L'analisi istologica rivela una complessa struttura a 7 strati, composta da diversi tipi di neuroni sui quali si scaricano informazioni provenienti dalle retine, dai nuclei della base, dalla corteccia e dagli organi di senso. In particolare, le informazioni visive si scaricano sugli strati superficiali, le informazioni acustiche sugli strati intermedi e le informazioni somatiche sugli strati profondi.
Analisi approfondite hanno rivelato una disposizione topografica dei neuroni del collicolo superiore; la mappa retinotopica superficiale è disposta in modo da essere congruente alla mappa somatosensitiva e uditiva presente negli strati più profondi. Non solo: negli strati di confine sono presenti neuroni multimodali ai quali scaricano contemporaneamente i neuroni della mappa visiva e i neuroni della mappa somatosensitava e uditiva. Il risultato è l'integrazione di diversi segnali sensoriali, fenomeno che trova riscontro nella percezione complessa dello spazio esplorato dai sensi.
Si immagini ad esempio di vedere una rondine in volo mentre emette il caratteristico canto: per opera delle integrazioni collicolari il sistema nervoso è in grado di associare il canto (un suono) alla rondine (l'immagine), oppure di dirigere lo sguardo verso il suono. Non solo: grazie alla precisa mappatura degli stimoli si ha la possibilità di associare un suono ad una data immagine e non ad un'altra. Ancora: una carezza inaspettata evocherà il puntamento istintivo della zona di cute toccata.
Si può allora considerare il collicolo superiore come un insieme di neuroni che nell'insieme formano un complesso sistema di trasduzione di coordinate. Dato che le coordinate naturali delle tre modalità sensoriali sono differenti (coordinate retiniche per la vista, coordinate centrate sul capo per l'udito, coordinate centrate sul tronco per la stimolazione propriocettiva), allora anche la rappresentazione spaziale degli stimoli avviene in modi differenti: le cellule visive propriamente dette codificano l'errore retinico, ovvero il vettore che rappresenta la distanza dello stimolo dalla fovea; le cellule quasi-visive (le cellule "in transizione" tra uno stato ed un altro) codificano la posizione degli stimoli in coordinate centrate sul capo; le cellule uditive sono sensibili sia alla posizione del suono che a quella degli occhi: sottraendo la posizione degli occhi alla posizione del suono codificata in coordinate centrate sul capo otteniamo la posizione del suono in coordinate centrate sulla retina.
Per quanto riguarda la trattazione per il solo apparato visivo, è interessante notare come le informazioni veicolate dal tratto ottico trovino nel collicolo superiore una stazione importante non solo per il confronto sensoriale ma anche per evocare una serie di movimenti oculari strettamente connessi con l'attività dei centri superiori.
Le informazioni provenienti dalla retina possono biforcarsi a livello del mesencefalo per dare una collaterale della via visiva centrale destinata al corpo genicolato laterale; inoltre esistono fibre che non si biforcano, destinate al solo (fibre tipo W) collicolo. Da qui le informazioni, dopo essere state adeguatamente integrate e confrontate, hanno diversi possibili destini:
- proiezione al pulvinar o al nucleo laterale posteriore del talamo e successivo smistamento[6] alla corteccia striata, prestriata, temporale media e parietale;
- proiezione pontino-bulbare ipsilaterale verso i neuroni oculomotori della formazione reticolare mesencefalica e la parte rostrale della formazione reticolare pontina;
- proiezione spinale verso i motoneuroni cervicali e toracici.
Queste proiezioni testimoniano i ruolo centrale del collicolo superiore nel controllo dei movimenti oculari e cervicali[7], finemente regolati da centri superiori come la corteccia e i nuclei della base. In particolare, zone diverse della corteccia controllano i movimenti oculari proiettando agli strati profondi del collicolo superiore. I nuclei della base, insieme alla regione media della corteccia parietale, sono i principali responsabili dell'attenzione visiva; la regione anteriore della corteccia parietale è invece connessa con i processi di fissazione.
Corpo genicolato laterale
[modifica | modifica wikitesto]Il corpo genicolato laterale (CGL) è una stazione talamica preposta allo smistamento delle informazioni visive provenienti dal tratto ottico. È opportuno ricordare che un singolo tratto ottico è composto dalle fibre provenienti da entrambi gli occhi. In particolare, dunque, un singolo corpo genicolato laterale riceve le afferenze dell'emiretina temporale ipsilaterale e della emiretina nasale controlaterale, cioè due rappresentazioni dell'emicampo visivo controlaterale. Inoltre, si deve ricordare che le fibre provenienti dalle singole retine sono composte da assoni provenienti sia dalle cellule gangliari di tipo M, sia da cellule gangliari di tipo P.
A livello del CGL questa divisione non solo si mantiene, ma viene ulteriormente scomposta per essere indirizzata a zone diverse della corteccia striata.
Il CGL è infatti diviso in 6 lamine concentriche; due di queste hanno afferenze esclusive dalle cellule gangliari M, mentre le restanti 4 ricevono da cellule gangliari P.
In particolare:
- lamina 1: cellule M, afferenze controlaterali;
- lamina 2: cellule M, afferenze ipsilaterali;
- lamina 3: cellule P, afferenze centro-off, ipsilaterali;
- lamina 4: cellule P, afferenze centro-off, controlaterali;
- lamina 5: cellule P, afferenze centro-on, ipsilaterali;
- lamina 6: cellule P, afferenze centro-on, controlaterali.
Si ha dunque che le lamine 1, 4 e 6 hanno afferenze miste controlaterali, mentre le lamine 2, 3 e 5 hanno afferenze miste ipsilaterali. Deve essere inoltre sottolineato che a livello del CGL si mantiene un preciso ordine retinotopico: la parte centrale del campo visivo è rappresentata posteriormente e centralmente, la parte periferica anteriormente, la metà inferiore medialmente e dorsalmente, la metà superiore lateralmente e ventralmente.
L'analisi in parallelo delle informazioni veicolate dalle cellule gangliari P ed M è particolarmente importante per la corretta percezione del colore e dell'illuminazione, oltre a fornire canali preferenziali per i processi di discriminazione spaziale e temporale.
| Caratteristiche dello stimolo | Sensibilità specifica cellule M | Sensibilità specifica cellule P |
|---|---|---|
| Contrasto di colore | No | Sì |
| Contrasto di luminanza | Elevato | Basso |
| Frequenza spaziale | Bassa | Elevata |
| Frequenza temporale | Elevata | Bassa |
La tabella qui sopra mostra le proprietà più importanti delle cellule M e P. Queste differenze verranno prese in considerazione successivamente per la trattazione della corteccia striata.

Radiazione ottica
[modifica | modifica wikitesto]Dal corpo genicolato laterale, l'informazione visiva viene proiettata alle aree di corteccia intorno alla scissura calcarina (corteccia visiva). L'insieme degli assoni tesi tra queste due strutture viene detto via genicolo-calcarina o radiazione ottica. La compartimentazione delle informazioni visive operata dalle varie strutture fa sì che la corteccia visiva "veda" l'emicampo visivo controlaterale.
Non solo; le informazioni provenienti dalla metà superiore del campo visivo vengono proiettate al labbro inferiore (rispetto alla scissura calcarina) della corteccia visiva, mentre le informazioni provenienti dalla metà inferiore del campo visivo vengono proiettate al labbro superiore.
La radiazione visiva assume quindi un aspetto a "ventaglio", con il manico posto nel corpo genicolato laterale e con il bordo perpendicolare alla scissura calcarina.
Questa considerazione è particolarmente importante in patologia, poiché permette di riconoscere il punto di lesione della via ottica a seconda del difetto di campo visivo riferito.
Corteccia visiva
[modifica | modifica wikitesto]Corteccia visiva primaria
[modifica | modifica wikitesto]È l'area 17 di Brodmann, nel lobo occipitale. Riceve le informazioni dalla parte controlaterale del campo visivo. Le innervazioni sono perfettamente retinotopiche, ovvero spazialmente organizzate come i recettori sulla retina. Dividendo il campo visivo in 4 settori le aree che ricevono la porzione in alto a sinistra si trovano sotto la scissura calcarina nell'emisfero destro, il settore in basso a destra giungerà sopra la scissura calcarina nell'emisfero sinistro.
La corteccia visiva è organizzata in 6 strati. Le afferenze arrivano principalmente al 4º strato (A, B, Cα, Cβ). Le fibre del cammino magno (M) arrivano allo strato 4Cα, da dove sono proiettate al 4B. Al 4Cβ e al 4A giungono le afferenze del cammino parvo, per essere inviate agli strati 2 e 3, mentre il cammino (K) giunge nei cosiddetti "blob". L'organizzazione in strati è fondamentale, non solo dal punto di vista anatomico ma anche funzionale. Infatti immersi in questi livelli vi sono due tipi principali di cellule, le cellule semplici (strati 4 e 6) e le cellule complesse (1, 2, 3 e 5).
In quest'area della corteccia si procede all'estrazione di bordi a partire dall'immagine in contrasto proveniente dal nucleo genicolato laterale. Ogni cellula semplice riceve in ingresso le informazioni di luminosità e le organizza in una zona centrale (eccitatoria) e una periferica (inibitoria) a formare un nuovo campo percettivo. Questo è organizzato in modo da permettere il riconoscimento di una linea opportunamente inclinata, posizionata nella zona eccitatoria. La funzione svolta, possiamo dire, si avvicina all'estrazione di bordi tipica dell'image processing. Le cellule complesse a loro volta hanno come ingresso l'informazione prodotta dalle cellule semplici. Ciascun campo percettivo viene affiancato ed in parte sovrapposto, a formare delle aree prive di una specifica zona di attivazione/inibizione, ma specifiche per il riconoscimento di bordi, secondo una specifica inclinazione, a prescindere dalla posizione nel campo percettivo. Le cellule dei blob invece sono specializzate nell'elaborazione dell'informazione cromatica, ma totalmente insensibili a direzioni.
Le cellule risultano quindi organizzate in colonne. Ogni colonna è formata dagli strati della corteccia, in ciascuno dei quali le cellule si attivano per una stessa direzione specifica con lo stesso campo percettivo come riferimento. Inoltre nelle zone adiacenti studi elettroencefalografici hanno dimostrato l'esistenza di colonne che codificano per le altre direzioni, ma per lo stesso campo percettivo. La sovrastruttura che comprende tutte le colonne di un determinato campo percettivo prende il nome di ipercolonna, ogni ipercolonna è affiancata a quella la cui afferenze provengono dall'area omologa dell'occhio controlaterale.
Le efferenze della V1 sono dirette sia ad altre zone corticali (dagli strati 2, 3 e 4B) sia ai centri profondi per le funzioni di feedback al nucleo genicolato laterale, al pretetto e al collicolo superiore (dagli strati 5 e 6).
Corteccia visiva secondaria
[modifica | modifica wikitesto]La corteccia visiva secondaria (V2), nota anche come pre-striata, è la seconda area più estesa della corteccia visiva.[8] Riceve informazioni dall'area V1 e si connette alla V3, V4 e V5. Invia anche forti proiezioni retroattive alla V1. La corteccia secondaria è divisa in quattro quadranti, che forniscono una mappa completa del campo visivo. Molti dei neuroni di quest'area sono regolati da caratteristiche visive semplici come l'orientamento, lo spazio, le dimensioni, il colore e la forma.[9][10]
Corteccia visiva terziaria
[modifica | modifica wikitesto]La corteccia visiva ha terziaria si trova di fronte alla secondaria. Può essere anatomicamente localizzata nell'area di Brodmann 19. L'area V3 svolgerebbe un ruolo nel processare i movimenti. La corteccia ventrale V3, ha connessioni più deboli con la l'area visiva primaria, e connessioni più forti con la corteccia temporale inferiore.
Corteccia cerebrale ed elaborazioni in parallelo
[modifica | modifica wikitesto]Galleria d'immagini
[modifica | modifica wikitesto]-
 Serie di immagini progressive che illustrano l'evoluzione dell'occhio.
Serie di immagini progressive che illustrano l'evoluzione dell'occhio. -
 Sezione del bulbo oculare: sono visibili, in particolare, la cornea, l'iride e il cristallino.
Sezione del bulbo oculare: sono visibili, in particolare, la cornea, l'iride e il cristallino. -
 Strati della retina.
Strati della retina. -
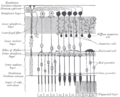 Anatomia microscopica della retina.
Anatomia microscopica della retina. -
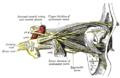 Innervazione di occhio e annessi, veduta laterale.
Innervazione di occhio e annessi, veduta laterale. -
 Innervazione di occhio e annessi, veduta dall'alto.
Innervazione di occhio e annessi, veduta dall'alto. -
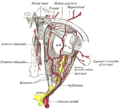 Vascolarizzazione di occhio e annessi, veduta dall'alto.
Vascolarizzazione di occhio e annessi, veduta dall'alto. -
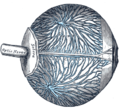 Sistema venoso "superficiale" dell'occhio.
Sistema venoso "superficiale" dell'occhio.








Note
[modifica | modifica wikitesto]- ^ Semir Zeki,La visione dall'interno. Arte e cervello, tit. or.Inner Vision.An Exploration of Art and the Brain, 1999, trad.it Paolo Pagli e Giovanna De Vivo, Torino, 2003, Bollati Boringhieri, pag 82, ISBN 88-339-1471-2
- ^ Semir Zeki, La visione dall'interno. Arte e cervello, tit. or.Inner Vision.An Exploration of Art and the Brain, 1999, trad.it Paolo Pagli e Giovanna De Vivo, Torino, 2003, Bollati Boringhieri, ISBN 88-339-1471-2, cap. 3 L'illusione di "vedere con gli occhi"
- ^ Eric R. Kandel, James H.Schwartz; Thomas M.Jessel, Analisi delle informazioni visive della retina, in Principi di Neuroscienze, Ristampa 2007, Milano, Casa Editrice Ambrosiana, 2003, p. 502, ISBN 88-408-1256-3.
- ^ In verità i nuclei oculomotori vengono preferenzialmente reclutati dal collicolo in stretta correlazione con gli "ordini" corticali e di altri nuclei per i movimenti di nistagmo fisiologico, di vergenza e di inseguimento lento, insieme ai movimenti saccadici, al riflesso optocinetico e al riflesso vestibolo-oculare, fenomeni trattati nell'analisi nervosa del movimento.
- ^ Nikos Logothetis, Jeffrey Schall,Neuronal correlates of subjective visual perception, 1989, in Science, 245, 4919, pp.761-763
- ^ Si noti che le fibre che proiettano dal collicolo superiore alla corteccia striata non fanno stazione nel corpo genicolato laterale, ma in altri nuclei posteriori del talamo.
- ^ Si noti che anche il pretetto è responsabile della genesi di riflessi di puntamento che coinvolgono la muscolatura cervicale. Questo è in accordo con la vicinanza di pretetto e collicolo superiore e con la difficoltà che si incontra nel definire con precisione i confini tra i due. Tuttavia, allo scopo di questa trattazione, si consiglia di considerare il collicolo superiore soprattutto per il controllo dei soli movimenti oculari
- ^ Gazzaniga, Michael S. e Mangun, G. R. (George Ronald), 1956-, Cognitive neuroscience : the biology of the mind, 3rd ed, W.W. Norton, 2009, ISBN 9780393927955, OCLC 225090391.
- ^ (EN) Jay Hegdé e David C. Van Essen, Selectivity for Complex Shapes in Primate Visual Area V2, in Journal of Neuroscience, vol. 20, n. 5, 1º marzo 2000, pp. RC61–RC61, DOI:10.1523/JNEUROSCI.20-05-j0001.2000. URL consultato il 6 maggio 2018.
- ^ (EN) Akiyuki Anzai, Xinmiao Peng e David C Van Essen, Neurons in monkey visual area V2 encode combinations of orientations, in Nature Neuroscience, vol. 10, n. 10, 16 settembre 2007, pp. 1313-1321, DOI:10.1038/nn1975. URL consultato il 6 maggio 2018.
Voci correlate
[modifica | modifica wikitesto]- Aree di Brodmann
- Cecità corticale
- Campo visivo
- Campi oculari supplementari
- Cervello
- Corteccia cerebrale
- Corteccia visiva
- Elettroretinografia
- Fovea
- Flusso ottico
- Illusione ottica
- Psicologia della Gestalt
- Occhio
- Oftalmologia
- Ottica
- Midriasi
- Nervo ottico
- Persistenza della visione
- Pretetto
- Potenziale d'azione
- Retina
- Talamo
- Tessuto nervoso
- Telencefalo
- Visione del colore
- Vista
Altri progetti
[modifica | modifica wikitesto] Wikimedia Commons contiene immagini o altri file su apparato visivo
Wikimedia Commons contiene immagini o altri file su apparato visivo
Collegamenti esterni
[modifica | modifica wikitesto]- Un mondo di Illusioni ottiche, su illuweb.it.
- Ottimo sito, ricco di informazioni sulla fisiologia, sulla patologia e sulle metodiche diagnostiche dell'apparato visivo, su occhio.it.
- (EN) Per localizzare la macchia cieca, su ophtasurf.com. URL consultato il 22 dicembre 2007 (archiviato dall'url originale il 12 dicembre 2007).
- (EN) Sito dell'università di Harvard sulla visione, su hubel.med.harvard.edu. URL consultato il 24 dicembre 2007 (archiviato dall'url originale il 25 dicembre 2007).
- (EN) Sito costantemente aggiornato sulle patologie e sulla fisiologia molecolare della retina, su retinacentral.org.
- (EN) Sito di ricerca sul sistema visivo dell'università dell'Utah, su webvision.med.utah.edu.
- (EN) Sito australiano di un gruppo di ricerca per le protesi oculari, su bionic.gsbme.unsw.edu.au. URL consultato il 24 dicembre 2007 (archiviato dall'url originale il 12 agosto 2011).
- (EN) Sito della clinica di ricerca del Dottor Schepens, massimo esperto mondiale della chirurgia dell'occhio, su schepens.harvard.edu.